TG101348 (Fedratinib)
Diphenyleneiodonium (chloride)
AMT (hydrochloride)
Aminoguanidine (hydrochloride)
作者:上海阿拉丁生化科技股份有限公司 2023-03-24T13:44 (访问量:5130)
TG101348 (Fedratinib)
Diphenyleneiodonium (chloride)
AMT (hydrochloride)
Aminoguanidine (hydrochloride)

程序性细胞死亡途径被先天免疫系统激活,以应对微生物感染和其他细胞应激源。焦亡、凋亡和坏死是三种程序性细胞死亡途径,已被广泛研究,并具有很好的特征。圣裘德儿童研究医院的Kanneganti小组的工作表明,这三种途径并不总是相互孤立地运作他们的工作描述了一种新的炎症程序性细胞死亡途径,PAN凋亡。之所以被称为PAN凋亡,是因为它涉及细胞死亡、凋亡和程序性坏死的一系列集体行为。
PAN细胞死亡可由多种细菌和病毒病原体诱导,包括单核增生李斯特菌、肠链球菌血清型鼠伤寒、水泡性口炎病毒(VSV)和甲型流感病毒(IAV)全视缺失也可能在重症COVID-19患者的炎症中发挥作用,在本文中后者是由严重急性呼吸综合征冠状病毒2 (SARS-CoV-2)引起的,我们综述了这种新描述的程序性细胞死亡途径的机制和调控,冠状病毒如何触发程序性细胞死亡,并讨论了最近的研究表明PAN细胞凋亡在covid -19相关炎症中的作用,为通过调节这种细胞死亡途径进行治疗干预提供了可能的机会。
PAN细胞凋亡由PAN凋亡体调节
PAN细胞凋亡的主调控因子是一种称为PAN凋亡体的多聚体细胞质蛋白复合体,由参与死亡、凋亡和程序性坏死的蛋白质组成。1,4它可以包括NOD-、LRR-和含吡啶结构域蛋白3 (NLRP3),含有CARD的凋亡相关斑点样蛋白(ASC),以及细胞凋亡蛋白酶,在死亡和炎症小体中起作用的蛋白质(图1)。凋亡蛋白半胱天冬酶-8抗原和坏死蛋白受体相互作用蛋白激酶1 (RIPK1)和RIPK3也可以被纳入PAN凋亡体。其他蛋白质成分包括半胱胺酸蛋白酶蛋白,作为支架,Z-DNA结合蛋白1 (ZBP1),作为先天免疫传感器,脂肪酸合成酶相关死亡结构域(FADD),作为适配器。通过PAN凋亡体的信号通路激活下游效应子gasdermin D (GSDMD), 半胱天冬酶-3和-7,以及混合谱系激酶结构域样(MLKL),分别执行细胞死亡, 凋亡和程序性坏死。
图1。ZBP1依赖的PAN凋亡体的激活在IAV感染时触发死亡、凋亡和坏死。
天然免疫传感器ZBP1是PAN凋亡体形成和激活的正向调节因子。1,4,5在IAV感染过程中,ZBP1识别病毒核糖核蛋白并诱导形成ZBP1依赖的泛体。依赖于ZBP1的泛光体由ZBP1(传感器)、RIPK3和RIPK1(坏死蛋白)、NLRP3、ASC、细胞凋亡蛋白酶-1(炎症小体/坏死蛋白)、细胞凋亡蛋白酶-8(凋亡蛋白)和支架细胞凋亡蛋白酶-6(图1)组成。该PAN凋亡体的形成导致RIPK3、细胞凋亡蛋白酶-8和NLRP3炎症小体的激活,导致PAN凋亡。
而TGF-β-活化激酶1 (TAK1)则是PAN细胞凋亡的负调控因子。TAK1的抑制与toll样受体(TLRs)或死亡受体(如TNF受体1 (TNFR1)、Fas、TRAIL-R或DR3)的信号耦合,促进ripk1依赖的PAN凋亡体的形成(图2)一种以这种方式诱导泛光症的细菌是耶尔森氏菌。耶尔森氏菌致病菌株可将效应蛋白YopJ分泌到巨噬细胞中抑制TAK1和NF-κB激酶抑制剂(IKK)。2,4这导致PAN凋亡的激活,细胞内病原体清除,以及包括IL-1β和IL-18在内的促炎细胞因子的释放。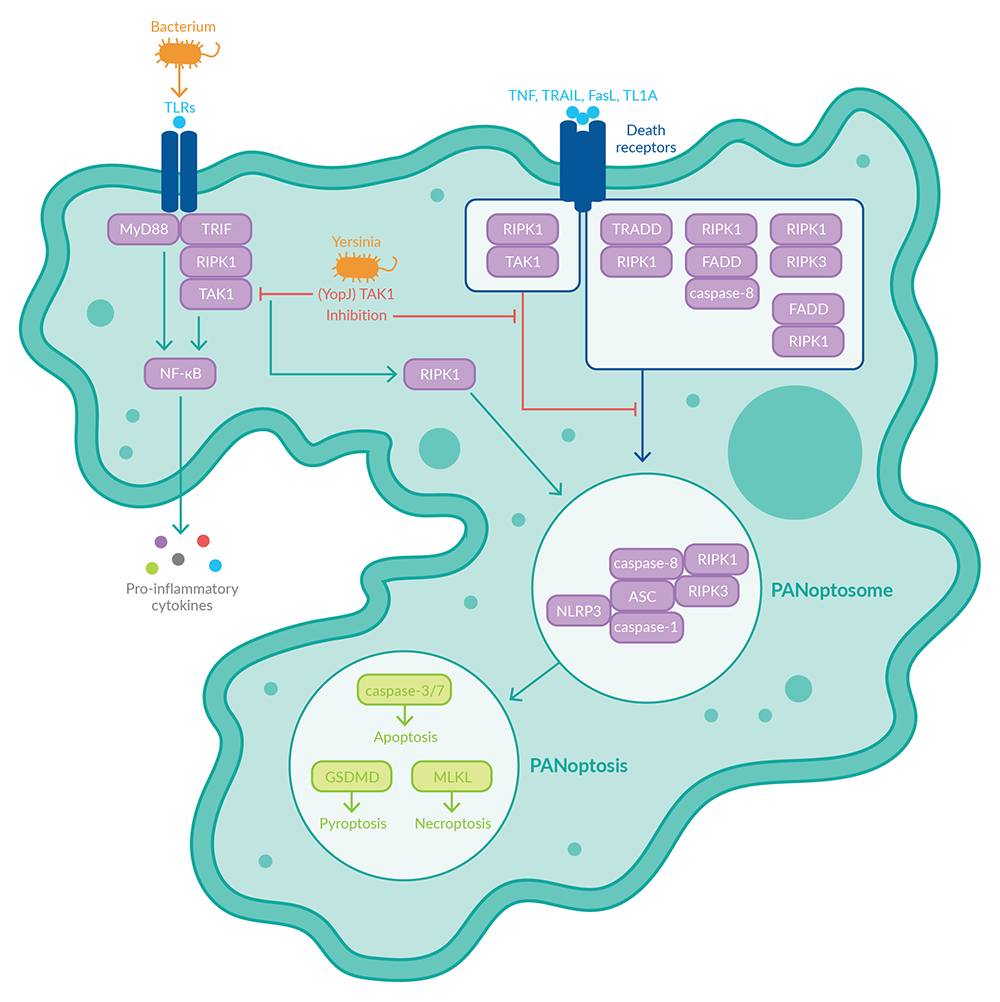
图2。致病性耶尔森氏菌通过抑制TAK1触发PAN凋亡,从而导致PAN凋亡体激活。
如上所述,IAV、VSV、单核增生L.和肠链球菌血清型鼠伤寒都能诱导巨噬细胞PAN细胞死亡仅抑制死亡、凋亡或坏死不足以保护巨噬细胞免受病原体诱导的细胞死亡。只有同时抑制这三种途径,例如通过删除编码细胞凋亡蛋白酶-1和细胞凋亡蛋白酶-11、细胞凋亡蛋白酶-8和坏死蛋白RIPK3的基因,才能保护巨噬细胞免受这些病原体诱导的PAN细胞死亡。这表明PAN凋亡的所有三个分支都是由细胞参与并介导细胞死亡以应对这些病原体。
冠状病毒激活程序性细胞死亡途径
乙型冠状病毒SARS-CoV、SARS-CoV-2、中东呼吸综合征冠状病毒(MERS-CoV)和小鼠肝炎病毒(MHV)已被证明可激活程序性细胞死亡途径(图3)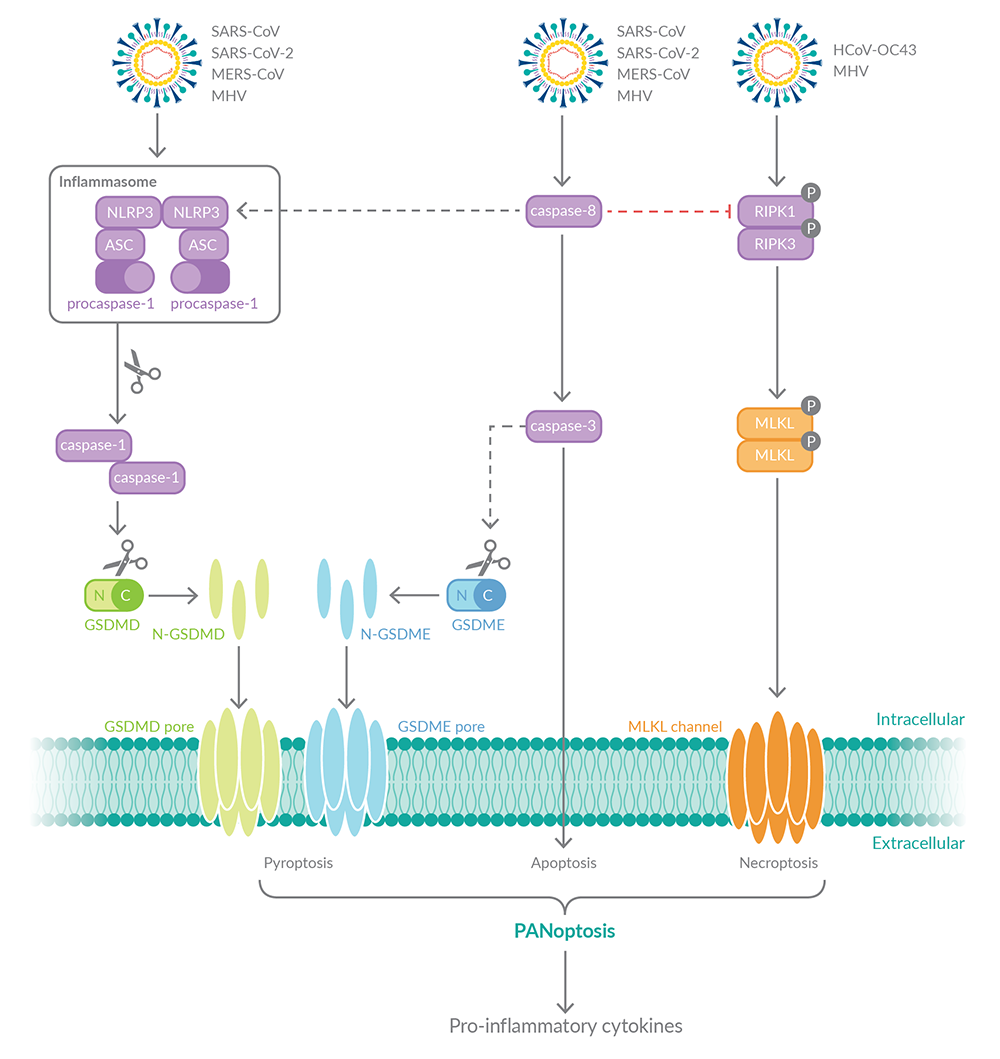
图3。冠状病毒激活多种程序性细胞死亡途径。
SARS-CoV、SARS-CoV-2和MERS-CoV均可致死亡和凋亡在SARS-CoV和SARS-CoV-2的情况下,这些病原体诱导的死亡伴随着促炎细胞因子IL-1β的分泌,以及NLRP3炎症小体的激活。HCoV-OC43是一种可导致普通感冒的冠状病毒,可诱导人类神经细胞坏死。还需要更多的研究来确定SARS-CoV、SARS-CoV-2和MERS-CoV是否也能诱导坏死。然而,小鼠冠状病毒MHV已被证明可以激活小鼠巨噬细胞中的所有三种泛光性细胞死亡途径,并且这种泛光性细胞死亡伴随着促炎细胞因子IL-1β、IL-18、IL-6和TNF.7的释放。
TNF-α和IFN-γ诱导PAN细胞死亡和类似COVID-19的炎症
Karki等人最近的研究表明,PAN凋亡可能在严重COVID-19.3患者的炎症反应中发挥作用。在小鼠中,促炎细胞因子TNF-α和IFN-γ的联合给药会导致死亡率增加,以及反映严重COVID-19患者观察到的各种表型,包括血清谷丙转氨酶(ALT)、天门冬氨酸转氨酶(AST)、血尿素氮(BUN)和铁蛋白水平升高。还有血小板减少和嗜中性粒细胞与淋巴细胞比例增加。在细胞中,TNF-α和IFN-γ诱导全视凋亡和细胞死亡。缺乏PAN凋亡体成分RIPK3和细胞凋亡蛋白酶-8的小鼠免受TNF-α-和IFN-γ-诱导的死亡。来自这些小鼠的巨噬细胞也受到TNF-α-和IFN-γ-诱导的全视和死亡的保护。这表明PAN凋亡是由TNF-α和IFN-γ联合诱导的,其病理与严重的COVID-19有相似之处。
作者接下来试图阐明TNF-α和IFN-γ.3诱导泛光症的信号通路他们发现JAK/STAT1/IRF1信号通路对调节很重要。在这一途径中,Janus激酶2 (JAK2)磷酸化JAK1,然后激活转录因子STAT1,诱导包括IFN调节因子1 (Irf1)在内的基因的转录(图4)。在TNF-α和IFN-γ处理的小鼠巨噬细胞中,JAK2和Irf1基因上调,以及在重症COVID-19患者中。通过删除Irf1或Stat1来破坏这一信号通路,可以保护小鼠巨噬细胞免受TNF-α-和IFN-γ-诱导的细胞死亡,以及在Irf1缺失的情况下诱导全视缺失。类似地,Stat1-/-小鼠免受TNF-α-和IFN-γ-诱导的死亡。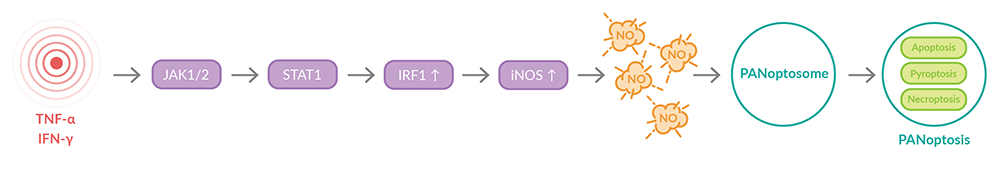
图4。TNF-α和IFN-γ诱导JAK/STAT1/IRF1信号通路和PAN凋亡。
Karki等人还证明,由JAK/STAT1/IRF1途径诱导的一氧化氮(NO)生成有助于TNF-α-和IFN-γ-诱导的PAN细胞死亡(图4)。3 IRF1 -/-小鼠巨噬细胞可降低诱导型一氧化氮合酶(iNOS)及其编码基因Nos2的表达。与此一致的是,与野生型细胞相比,当TNF-α和IFN-γ刺激时,Irf1-/-和Stat1-/-细胞都减少了NO的产生。通过删除Nos2或使用NO生成抑制剂L-NAME或1400W来干扰NO生成,可以保护细胞免受TNF-α-和IFN-γ-诱导的细胞死亡。
阿拉丁提供了多种工具来研究PAN凋亡和其他程序性细胞死亡途径,包括PAN凋亡体成分和调节剂的小分子抑制剂,其他细胞死亡途径,JAK/STAT信号,和诱导性一氧化氮合酶产生。
PAN凋亡体抑制剂组成与调节蛋白 | |||
PAN凋亡体抑制剂组成/调节蛋白 | 抑制剂 | PAN凋亡体抑制剂组成/调节蛋白 | 抑制剂 |
NLRP3 | NLRP3i | RIPK3/RIP3 Kinase | GSK872 |
MCC950 | RIPK1/RIP1 Kinase | (±)-Necrostatin-2 | |
Dapansutrile | Necrostatin-1 | ||
INF39 | Necrostatin-5 | ||
Caspase-1 | Ac-YVAD-CMK | GSK2982772 | |
Ac-YVAD-CHO | GSK481 | ||
VX-765 | TAK1 | (5Z)-7-Oxozeaenol | |
Caspase-8 | Ac-IETD-CHO (trifluoroacetate salt) | Takinib | |
Caspase-6 | Ac-VEID-CHO (trifluoroacetate salt) |
| |
其余与细胞死亡、凋亡和坏死有关的抑制剂 | |||
靶标蛋白 | 抑制剂 | 靶标蛋白 | 抑制剂 |
Pan-细胞凋亡蛋白酶 | Z-VAD(OMe)-FMK | 细胞凋亡蛋白酶-3 | Z-DEVD-FMK |
Q-VD-OPH | Ac-DEVD-CMK | ||
Z-VAD(OH)-FMK | Caspase-3 Inhibitor VII | ||
Emricasan | 细胞凋亡蛋白酶-1/3 | Z-YVAD-CMK (trifluoroacetate salt) | |
Boc-D-FMK | MLKL | Necrosulfonamide | |
Z-Asp-CH2-DCB | 程序性死亡 | Necrostatin-2 | |
细胞凋亡蛋白酶-3/7 | Ac-DEVD-CHO | Necrostatin-7 | |
Caspase-3/7 Inhibitor I |
|
| |
JAK 抑制剂 | iNOS 抑制剂 |
Ruxolitinib | L-NAME (hydrochloride) |
Baricitinib | 1400W (hydrochloride) |
Filgotinib | L-NIL (hydrochloride)
上海阿拉丁生化科技股份有限公司 商家主页地 址: 上海市新金桥路36号财富中心16楼 联系人: 阿博士 电 话: 400-620-6333 传 真: 021-50323701 Email:market@aladdin-e.com 相关咨询【阿拉丁】系统学分类 | 理清结构归属,秒得同类产品 (暂无发布时间 浏览数:8743) 【阿拉丁】免疫沉淀通用实验方案 (2025-06-20T00:00 浏览数:32148) 【阿拉丁】蛋白质印迹的二抗选择指南 (2025-04-18T00:00 浏览数:35259) 阿拉丁抗菌配体 | 高效靶向,助力抗感染药物研发 (2025-03-27T00:00 浏览数:38984) 【阿拉丁】哺乳动物细胞冷冻保存方案 (2025-03-14T00:00 浏览数:38485) 【阿拉丁】新春开学季,满减钜惠 (2025-03-07T00:00 浏览数:35237) 【阿拉丁】固定免疫沉淀实验方案 (2025-02-13T00:00 浏览数:43444) 【阿拉丁】细胞培养通用指南 (2025-02-05T00:00 浏览数:42348) 阿拉丁蛋白Marker | 三色预染,分子量标记新标准 (2025-01-23T00:00 浏览数:43963) 阿拉丁®内参抗体 (2025-01-15T00:00 浏览数:43879) ADVERTISEMENT
|